
Olá a todos, estou ressuscitando o blog depois de muito tempo. Agora como professor da UFRN. Estou também inaugurando a publicação de textos dos alunos do curso de graduação "Ecologia, Evolução e Desenvolvimento". A primeira é a aluna Gabriela Neres de Oliveira e Silva.
 |
Muitas espécies de borboletas
são conhecidas pelos padrões observados nas asas. Além da beleza incontestável,
os padrões das borboletas podem ter diversas funções ecológicas, como
possibilitar a termorregulação do inseto, camuflagem e mimetismo, quando o
animal finge ser um outro (mais perigoso por exemplo) para seu benefício
ecológico. Entretanto, pouco se sabe como estas características surgem. Em
muitas espécies com dimorfismo entre sexos, os padrões das asas podem ser
decisivos para a seleção sexual. Seleção sexual pode ser entendida como uma
competição entre indivíduos de uma espécie para a realização da cópula. Assim, existe
uma pressão pelo desenvolvimento de caracteres que confiram aos indivíduos
maior sucesso em copular. Estes fenótipos podem, por exemplo, indicar aos
parceiros que o acasalamento com aquele indivíduo terá uma vantagem. Esse
mecanismo pode selecionar comportamentos sociais das espécies². Mas na maioria
das borboletas, ainda se desconhece qual o mecanismo de evolução e manutenção
desses padrões das asas. A borboleta que falarei hoje transmite sinais visuais
que atrairão ou não os machos dependendo do que come quando lagarta.
Como os padrões surgem nas asas das borboletas?
Fora todas as questões químicas e físicas dos pigmentos, o processo de síntese
dos pigmentos ocorre durante o estágio de pupa do inseto. Além das
características gênicas das espécies, outro fator importante nesse processo é a
dieta da larva. As larvas de borboletas não se alimentam de qualquer planta (ou
mesmo nem de plantas), e essa relação forte entre determinada espécie de
borboleta e espécie de planta é essencial e até indica uma coevolução. As larvas
de borboletas azuis (Lycaenidae), por
exemplo, absorvem e armazenam flavonoides das plantas que se alimentam. Durante
a síntese de pigmentos na pupa, esses flavonoides são então transferidos para
as asas. Flavonoides são compostos encontrados principalmente em flores, botões
de flores e frutas jovens. Todos eles absorvem ultravioleta e, dependendo do
tipo, podem ter cores que conseguimos enxergar. Observe na figura abaixo a
proximidade do espectro de luz ultravioleta e o da luz visível por nós.
 |
| Peter Hermes Furian/ shutterstock.com |
Onde eu estou querendo chegar com toda essa
explicação? Dois pesquisadores buscaram compreender qual a função ecológica da
absorção de flavonoides em Polyommatus icarus,
uma espécie de borboleta azul em que mais se estudou essa absorção. Ao saber
que a presença dos flavonoides mudava as cores das asas, eles se perguntaram se
isso transmite algum sinal entre indivíduos da mesma espécie e qual informação
esse sinal transmite.
A nossa P.
Icarus apresenta um notável dimorfismo sexual (veja figura abaixo), em que o macho é
azul na parte dorsal e a fêmea apresenta uma cor marrom escuro. Na parte
ventral, as cores são mais próximas, sem tanta diferenciação. No que diz respeito ao comportamento de corte,
depois que os machos localizam uma fêmea eles se aproximam e podem usualmente
agitarem-se ao redor dela, até pousar e tentar copular. Pode acontecer de o
macho desistir no meio do “ritual” e a fêmea ainda pode recusar o acasalamento.
Logo ambos os sexos apresentam essa escolha de parceiro.
 |
| P. Icarus: A. Espécime macho e B. um espécime fêmea. |
O estudo foi focado no comportamento de corte
dos machos com as fêmeas. As larvas foram criadas em duas dietas, ambas com a
mesma base alimentar, só que uma dieta com adição de flavonoide que a larva absorve
(borboleta com flavonoide) e outra não. Após chegarem à fase adulta, as
borboletas fêmeas foram mortas por congelamento e desidratadas em uma sala para
serem usadas como modelos de atração dos machos. Com isso, eles compararam o reflexo e o
espectro de luz das asas de cada tipo de borboleta, tanto na visão dorsal
quanto ventral, procurando compreender melhor as diferenças nas questões de
ondas luminosas. Para olhos humanos, as asas podem apresentar nenhuma
diferença, mas é possível notar com os dados coletados que o reflexo da luz ultravioleta,
que não é visível para nós, é muito maior na borboleta sem flavonoide (veja na figura abaixo).
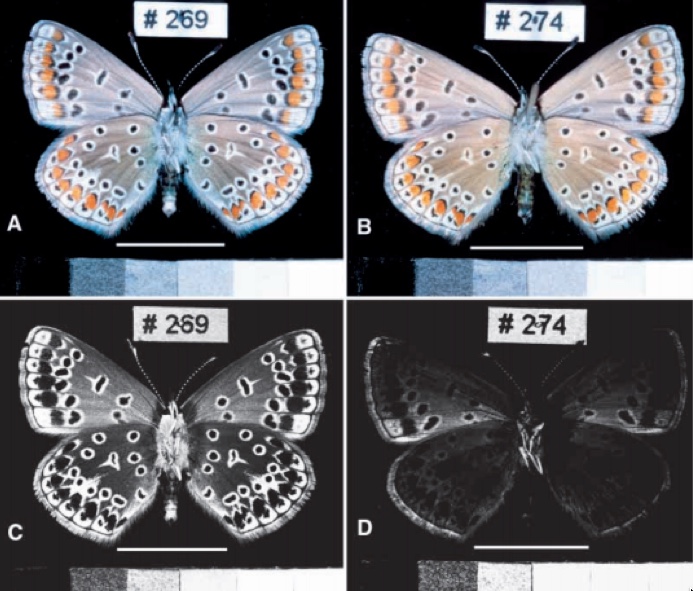 |
| Padrão de coloração das asas das fêmeas. A,B. Fotografias nos comprmentos de onda visíveis aos nossos olhos. C,D. Fotografias em comprimentos de onda U.V. . A coluna da esquerda mostra um animal controle (sem flavonoides na dieta) e a coluna da direita uma animal que absorveu flavonoides e absorve muito mais U.V. em suas asas (ao invés de refletir como o animal controle). |
Mostrou-se ainda que apenas as manchas brancas (e pouco da cor de fundo da asa)
na parte ventral têm uma grande diferença de reflexão da luz, sendo a borboleta
sem flavonoide que mais reflete. Os pesquisadores ainda simularam
fotorreceptores de borboletas, sendo eles do azul, verde, vermelho e também
ultravioleta. No fotorreceptor de ultravioleta obteve-se diferenças de valores
evidentes entre as cores da parte ventral da asa do inseto.
Dentre as variadas conclusões, pode-se destacar
que as larvas de borboleta crescem melhor quando se alimentam de flores e
botões, partes da planta com flavonoides que são absorvidos, e se alimentando
dessas partes as relações com formigas podem acontecer e ser favoráveis às
lagartas, pois as formigas ficam com as folhas e as lagartas com as flores, não
acontecendo competição. Por isso, a incorporação de flavonoides pode ser uma
forma de indicar uma boa alimentação na fase jovem do inseto. Logo, demonstra um
parceiro de melhor qualidade e isso pode ser indicado nos padrões das asas.
Ademais, o tamanho do corpo pode indicar uma fêmea mais fértil e as fêmeas com
flavonoides apresentam uma massa maior. Este trabalho mostrou que a atração é
influenciada pelos fatores visuais e principalmente relacionada à superfície
ventral nas asas.
Em suma, os flavonoides absorvidos pelas
borboletas durante o estágio larval podem mudar claramente os padrões das asas
e isso influencia na escolha dos machos para a cópula. Os efeitos dos
flavonoides nas cores quase sempre ocorrem com elas alcançando até ondas
ultravioletas. Apesar de serem necessárias mais bases teóricas e matemáticas para
estudar essas ondas é possível observar o impacto dessas interações da larva e
seu ambiente, mais especificamente seu alimento, na ecologia, relações
intraespecíficas, e até podendo refletir em aspectos para a evolução da
espécie.
Referências
KNÜTTEL, Helge; FIEDLER, Konrad. Host-plant-derived
variation in ultraviolet wing patterns influences mate selection by male
butterflies. Journal Of Experimental
Biology, Cambridge, v. 204, n. 14, p.2447-2459, 19 abr. 2001.